Summary: The effects of Garcinia kola (G. kola) seed extract on oestrous cycle, ovulation and foetal development were studied in adult female Sprague-Dawley (S-D) rats. Cyclic female rats weighing 150 to 200g were divided into three experimental groups and a control group. Group 1 was fed with 200mg/kg body weight of theextract on proestrous. Group 2 received 200mg/kg body weight of the extract daily for six weeks, while group 3, consisted of pregnant rats which received the same dose of the extract on days 1-5, 7-9th, 13th and 14th day of gestation. In groups 1 and 2, vaginal lavage was taken daily to monitor the oestrous cycle and ovulation. In group 3, gestational parameters monitored were number of total implants, resorption and dead foetuses. Live foetuses were weighed and examined for external malformation and variation. The results showed that the oestrous cycle was altered for the first two weeks after commencement of extract but returned to normal from the third week. This was indicated by the irregular pattern of oestrous with a prolonged dioestrus observed in the treated rats. Ovulation was partially blocked as shown by the reduced number of ova observed in the oviduct from the treated rats compared with control (p < 0.05). There was a significant decrease in the weight of foetuses from the treated rats (p < 0.05) while 7% of the foetuses from pregnant rats, which received treatment for the first five days of gestation, had malformed left upper limb. Results suggest that G. kola seed at 200mg/kg body weight administered alters oestrous cycle in rats, partly inhibits ovulation and may produce duration dependent teratogenicity in foetal rats.
Key Words: Garcinia kola, ovulation, oestrous cycle, foetal development.
Introduction
The quest for naturally occurring compounds of herbal or plant origin that could be of benefit as contraceptive and fertility control agents, stimulated our interest inGarcinia kola Heckel (Guttiferae) seeds (‘bitter kola’) which is widely consumed as a stimulant (Atawodi et al., 1995). Traditional African medicinal uses include, treatment of cough, purgative, anti-parasitic, anti-microbial( Madubunyi, 1995, Okunji et al.,1995; Adefule-Ositelu et al.,2004) The seed is used in the treatment of diarrhoea (Braide, 1991) bronchitis and throat infections(Adesina et al., 1995; Orie and Ekon, 1993) and liver disorders (Iwu et al., 1990). The seed, Garcinia kolaenjoys a folk reputation in Africa as a poison antidote (Kabangu et al.,1987). In addition, the plant possesses anti-hepatotoxic (Braide, 1991; Akintowa and Essien, 1990), antioxidant (Olatunde et al., 2004), hypoglycemic (Iwu et al., 1990; Odeigah et al.,1999) and aphrodisiac properties (Ajibola and Satake, 1992). Garcinia kolaseeds contain biflavonoid (Kolaviron) capable of having anti-inflammatory properties (Braide,1993) and is a natural antioxidant (Olatunde et al., 2002, Terashima et al., 2002) . Constituents of the seed of G. kola include 1-3, 8-11 benzophenones, Garcinia biflavonones (GB-1, GB-2) and kolaflavonone (Cotterih et al., 1978). Apigenin based flavonoids represent 60% of the total flavonoids present in the diethyl ether fraction of G. kola seeds (Iwu and Igboko,1982). The biological activities of flavonoids include action against allergies, inflammation, free radicals, hepatoxins (Terashima et al., 2002). The anti-inflammatory properties of G. kola seeds made us investigate the effect of the seed extract on ovulation and the oestrous cycle to assess its contraceptive effects. The importance of the anti-inflammatory property of G. kola is emphasized by evidence that ovulation, an important process in female reproductive function is believed to be an inflammatory process (Epsey, 1980; Epsey, 1994). Ovulation in rat is brought about by a luteinising hormone (LH) surge. The circulating levels of LH begins to rise on the afternoon of prooestrus, about 2pm to 3pm and reaches peak at about 5-7pm. This rapid surge induces follicular rupture and ovulation. Ovulation can be blocked experimentally by high doses of antiinflammatory drugs administered before the LH surge because once the levels start to rise it may not be brought down by any drug (Gaytan et al., 2002).
Materials and Methods48 adult female Sprague-Dawley rats were collected from the Animal house of the College of Medicine, University of Lagos. They were acclamatized for 2 weeks in the rat control room under standard conditions of temperature and illumination (12 hours dark: 12 hours light) cycle. They were fed commercially available rat’s pellets (Ladokun feeds, Ibadan) and had access to drinking water ad libitum. Ratsthat underwent 2 successive 4 or 5-day cycles, weighing 150 - 200g were used. They were divided into subgroups of five each.
The plant material, G. kola seeds were bought from the local market in Lagos. The outer coats were removed and the seeds cut into pieces and air-dried. The dried seeds were ground to fine powder and extraction done using 70% alcohol in a Soxhlet extraction. The yield was concentrated by evaporation in a water bath and dried to solid form. 2.0g of the extract was measured out and dissolved in 100mls of distilled water to give 20mg/ml.
The animals were divided into three experimental groups each with a control group. In the ovulation experiment, group I (n=20), animals were replicated into control and three experimental subgroups a, b, c; of 5 rats each which received the extract, 200mg/kg body weight at 10am, 2pm and 6pm respectively. The control group received distilled water. Vaginal smears were obtained daily by vaginal lavage to monitor ovulation and oestrous cycle. At the end of group 1 experiment, the animals were sacrificed using ether anesthesia and the fimbriated part of the oviduct was dissected out from the rats, suspended in normal saline and placed on a microscopic slide with a cover slip to count the number of ova in the oviduct.
In the oestrous cycle experiments, group II (n=6) the animals received 200mg/kg body weight of G. kola extract once daily for six weeks. The pattern of oestrous cycle before and after administration of extract was studied and the animals served as their control. In the teratogenic experiment, group III (n=22) the animals were mated during the proestrous to oestrus night and the presence of spermatozoa was determined by microscopic examination of the vaginal smear the next morning. The presence of spermatozoa indicated conception and represented day 1 of pregnancy (Oderinde et al., 2002). These pregnant rats were subdivided into groups a, b, and c which received 200mg/kg body weight of G. kola on days 1 to 5 of gestation (implantation studies) for group IIIa, 7-9 days of gestation for IIIb (beginning of organogenesis), and 14th and 15th day of gestation for group IIIc. The control group ‘d’ received distilled water. Body weight, food consumption, gross appearance and behaviour were monitored daily. On day 21 of gestation, foetuses were removed from pregnant rats by ventral laparatomy and examined. The number of total implants, resorption, live and dead foetuses recorded. Live foetuses were removed from the uterus and weighed, and examined for gross malformations. Foetal parameters such as foetal number, weight, crown-rump-length, and length of umbilical cord, and placental weight were measured.
Statistics: Results were expressed as Mean ± SEM. For experiments 1 and 3, ANOVA with one degree of classification was used for comparison of more than two means followed by Scheffe’s post hoc test. P values of 0.05 or less were considered significant (Table 1 and 3). Simple percentages was used to present the degree of alteration in different phases of oestrous cycle in experiment 2 (Table 2).
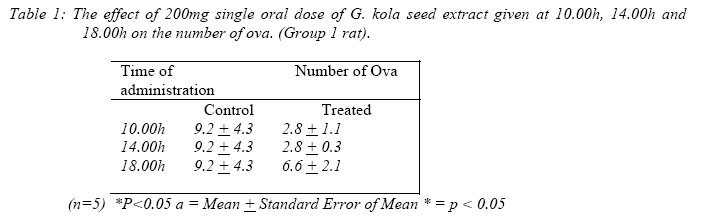
ResultsOvulation: The number of ova in the oviduct of treated rats was significantly reduced after commencement of treatment (p<0.05) when compared with the control with values of 9.2 ± 4.3 for control and 2.8 ± 1.1 for the treated at 10am and values of 9.2 ± 4.3 for control, 2.8 ± 0.3 for the treated at 2pm. There was no significant difference in the number of ova in treated rats fed at 6pm (Table 1)
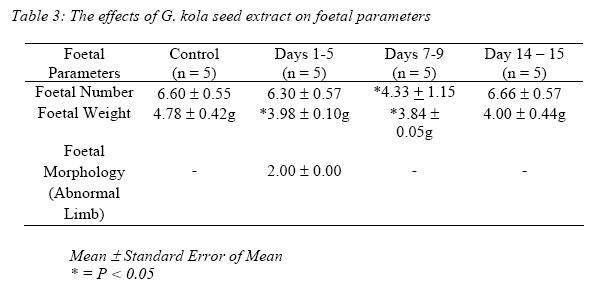
Oestrus Cycle: The normal pattern of oestrous cycle was significantly altered (99.2 – 35.8%) in the treated rats after two weeks (three cycles) of extract administration but returned to normal later to normal (90.9%) (Table 2). It was observed that cycles in the last four weeks were similar to control values (six cycles). The duration of dioestrous phase was increased from 26.29% to 41.0% in the treated rats after two weeks while the duration of proestrous and oestrous phases were reduced from 16.9% to 12.9% and 20.0% to 11.5% respectively.
Materials and Methods48 adult female Sprague-Dawley rats were collected from the Animal house of the College of Medicine, University of Lagos. They were acclamatized for 2 weeks in the rat control room under standard conditions of temperature and illumination (12 hours dark: 12 hours light) cycle. They were fed commercially available rat’s pellets (Ladokun feeds, Ibadan) and had access to drinking water ad libitum. Ratsthat underwent 2 successive 4 or 5-day cycles, weighing 150 - 200g were used. They were divided into subgroups of five each.
The plant material, G. kola seeds were bought from the local market in Lagos. The outer coats were removed and the seeds cut into pieces and air-dried. The dried seeds were ground to fine powder and extraction done using 70% alcohol in a Soxhlet extraction. The yield was concentrated by evaporation in a water bath and dried to solid form. 2.0g of the extract was measured out and dissolved in 100mls of distilled water to give 20mg/ml.
The animals were divided into three experimental groups each with a control group. In the ovulation experiment, group I (n=20), animals were replicated into control and three experimental subgroups a, b, c; of 5 rats each which received the extract, 200mg/kg body weight at 10am, 2pm and 6pm respectively. The control group received distilled water. Vaginal smears were obtained daily by vaginal lavage to monitor ovulation and oestrous cycle. At the end of group 1 experiment, the animals were sacrificed using ether anesthesia and the fimbriated part of the oviduct was dissected out from the rats, suspended in normal saline and placed on a microscopic slide with a cover slip to count the number of ova in the oviduct.
In the oestrous cycle experiments, group II (n=6) the animals received 200mg/kg body weight of G. kola extract once daily for six weeks. The pattern of oestrous cycle before and after administration of extract was studied and the animals served as their control. In the teratogenic experiment, group III (n=22) the animals were mated during the proestrous to oestrus night and the presence of spermatozoa was determined by microscopic examination of the vaginal smear the next morning. The presence of spermatozoa indicated conception and represented day 1 of pregnancy (Oderinde et al., 2002). These pregnant rats were subdivided into groups a, b, and c which received 200mg/kg body weight of G. kola on days 1 to 5 of gestation (implantation studies) for group IIIa, 7-9 days of gestation for IIIb (beginning of organogenesis), and 14th and 15th day of gestation for group IIIc. The control group ‘d’ received distilled water. Body weight, food consumption, gross appearance and behaviour were monitored daily. On day 21 of gestation, foetuses were removed from pregnant rats by ventral laparatomy and examined. The number of total implants, resorption, live and dead foetuses recorded. Live foetuses were removed from the uterus and weighed, and examined for gross malformations. Foetal parameters such as foetal number, weight, crown-rump-length, and length of umbilical cord, and placental weight were measured.
Statistics: Results were expressed as Mean ± SEM. For experiments 1 and 3, ANOVA with one degree of classification was used for comparison of more than two means followed by Scheffe’s post hoc test. P values of 0.05 or less were considered significant (Table 1 and 3). Simple percentages was used to present the degree of alteration in different phases of oestrous cycle in experiment 2 (Table 2).
ResultsOvulation: The number of ova in the oviduct of treated rats was significantly reduced after commencement of treatment (p<0.05) when compared with the control with values of 9.2 ± 4.3 for control and 2.8 ± 1.1 for the treated at 10am and values of 9.2 ± 4.3 for control, 2.8 ± 0.3 for the treated at 2pm. There was no significant difference in the number of ova in treated rats fed at 6pm (Table 1)
Oestrus Cycle: The normal pattern of oestrous cycle was significantly altered (99.2 – 35.8%) in the treated rats after two weeks (three cycles) of extract administration but returned to normal later to normal (90.9%) (Table 2). It was observed that cycles in the last four weeks were similar to control values (six cycles). The duration of dioestrous phase was increased from 26.29% to 41.0% in the treated rats after two weeks while the duration of proestrous and oestrous phases were reduced from 16.9% to 12.9% and 20.0% to 11.5% respectively.
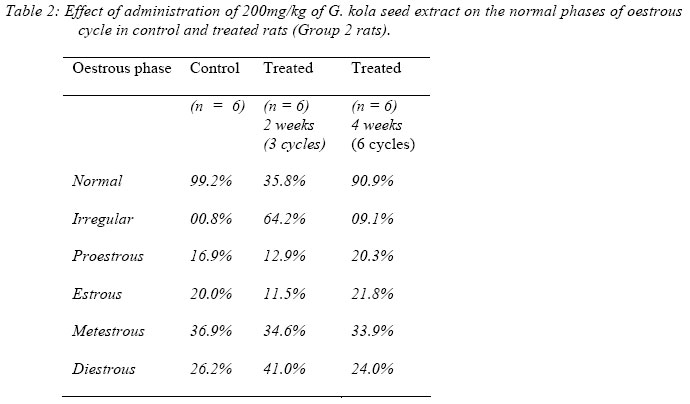
Gestational Parameters and Morphologic Defects: All dams on study survived to their scheduled termination day. There were no abortions, no early deliveries and no death of animal during the study. Data on rat foetal weight are presented in Table 3. The weights of foetuses produced by pregnant rats, which received G. kolaseed extract were significantly reduced (p<0.05). Among the experimental groups, 7% of the foetuses from pregnant rats fed on the first five days of gestation showed morphological anomalies (truncated limbs). Parameters for growth (crown-rump, placenta weight) were not affected. There were no resorption and no post-implantation sites.
DiscussionG. kola seeds have been reported to have an anti-inflammatory activity (Madubunyi, 1995; Braide,1993). These studies have revealed that the process of ovulation is comparable to an inflammatory process (Epsey, 1980). Anti-inflammatory drugs have been employed in blocking ovulation (Gaytan et al., 2002). The anti-inflammatory property of G. kola seed may be responsible for its observed effect in blocking ovulation when administered to the rats before the surge of lutenising hormone (which causes follicular rupture and release of ova) reached its peak (Freeman,1988). The anti-inflammatory property of flavonoids is believed to result from inhibition of cyclo-oxygenase enzyme (Liang et al., 1999). This implies that the mechanism of the anti-ovulatory property may be similar to that induced by indomethacin, a potent cyclo-oxygenase enzyme inhibitor (Epsey, 1983) suggesting that G. kola may not inhibit ovum maturation but causes abnormal follicle rupture (Gaytan et al., 2002).
Cyclo-oxygenase, which converts arachidonic acid derived from cell membranes to prostaglandins (PG), has two isomers, cyclooxygenase-1 (COX-1) and cyclooxygenase-2 (COX-2) (Osau et al.,2001). COX-1 is the endogenous form of the enzyme necessary for production of PG while COX-2 is thought of as being an inducible enzyme associated with inflammation. The latter is thought to be essential for the ovulatory mechanism. It was revealed that all traditional non-steroidal anti-inflammatory drugs affect the action of both COX-1 and COX-2 but produce most of their effects by blocking COX-2 (Staud, 2000). COX-2 is induced in various cells by stimulation of cytokines and/or growth factors. It is expressed in many conditions and organs such as in acute inflammation, bone resorption, kidneys, brain and the female reproductive organs (Katori and Majima, 2000). COX-2 deficient mice suffer from defect in reproductive functions such as ovulation and fertilization (Lim et al., 1997), implying that COX- 2 is important in ovulation. These reports (Katima and Majima, 2000;Lim et al., 1997) indicate that COX-2 is an essential enzyme for follicular rupture through metabolites of arachidonic acids, which play important roles in follicular rupture through the activation of proteases, neo-vascularization, leukocyte migration and smooth muscle contraction (Katori and Majima, 2000). The results of our study suggest that G. kola seed may block ovulation by inhibiting cyclooxygenase activity (perhaps COX-2) and PG Synthesis.
Some flavonoids (including apigenin based) suppress the formation of COX-2 thus playing an important role in the prevention of cancer and inflammation, partly via inhibiting COX-2 enzymes. This property is also currently under trial in chemoprevention potentials against human cancers as many types of cancer cells use COX-2 to propagate (Liang et al., 1999). It has been reported that daily consumption of large amount of quercetin or apigenin rich food may not be effective in inhibiting cyclooxygenase activity or platelet aggregation in human volunteers (Janssen et al., 1998)
Janssen et al., (1998) in ‘effects of flavonoid on hemostasis: Results from in vitro and a dietary supplement study wrote that only high concentrations of these flavonoids about 2500 µmol/L, which cannot be attained in-vivo by dietary consumption, inhibit collagen-induced aggregation in vitro. From their data, peak apigenin concentration in human plasma was < 1.1 µmol/L. The concentration we administered may have been enough to inhibit cyclooxygenase in relation to ovulation.
In conclusion,these studies suggest that administration of G. kola seed extract may partially block ovulation, alters oestrous cycle with a prolonged dioestrous, and may cause a dose dependent adverse effect on foetal development in S-D rats.
DiscussionG. kola seeds have been reported to have an anti-inflammatory activity (Madubunyi, 1995; Braide,1993). These studies have revealed that the process of ovulation is comparable to an inflammatory process (Epsey, 1980). Anti-inflammatory drugs have been employed in blocking ovulation (Gaytan et al., 2002). The anti-inflammatory property of G. kola seed may be responsible for its observed effect in blocking ovulation when administered to the rats before the surge of lutenising hormone (which causes follicular rupture and release of ova) reached its peak (Freeman,1988). The anti-inflammatory property of flavonoids is believed to result from inhibition of cyclo-oxygenase enzyme (Liang et al., 1999). This implies that the mechanism of the anti-ovulatory property may be similar to that induced by indomethacin, a potent cyclo-oxygenase enzyme inhibitor (Epsey, 1983) suggesting that G. kola may not inhibit ovum maturation but causes abnormal follicle rupture (Gaytan et al., 2002).
Cyclo-oxygenase, which converts arachidonic acid derived from cell membranes to prostaglandins (PG), has two isomers, cyclooxygenase-1 (COX-1) and cyclooxygenase-2 (COX-2) (Osau et al.,2001). COX-1 is the endogenous form of the enzyme necessary for production of PG while COX-2 is thought of as being an inducible enzyme associated with inflammation. The latter is thought to be essential for the ovulatory mechanism. It was revealed that all traditional non-steroidal anti-inflammatory drugs affect the action of both COX-1 and COX-2 but produce most of their effects by blocking COX-2 (Staud, 2000). COX-2 is induced in various cells by stimulation of cytokines and/or growth factors. It is expressed in many conditions and organs such as in acute inflammation, bone resorption, kidneys, brain and the female reproductive organs (Katori and Majima, 2000). COX-2 deficient mice suffer from defect in reproductive functions such as ovulation and fertilization (Lim et al., 1997), implying that COX- 2 is important in ovulation. These reports (Katima and Majima, 2000;Lim et al., 1997) indicate that COX-2 is an essential enzyme for follicular rupture through metabolites of arachidonic acids, which play important roles in follicular rupture through the activation of proteases, neo-vascularization, leukocyte migration and smooth muscle contraction (Katori and Majima, 2000). The results of our study suggest that G. kola seed may block ovulation by inhibiting cyclooxygenase activity (perhaps COX-2) and PG Synthesis.
Some flavonoids (including apigenin based) suppress the formation of COX-2 thus playing an important role in the prevention of cancer and inflammation, partly via inhibiting COX-2 enzymes. This property is also currently under trial in chemoprevention potentials against human cancers as many types of cancer cells use COX-2 to propagate (Liang et al., 1999). It has been reported that daily consumption of large amount of quercetin or apigenin rich food may not be effective in inhibiting cyclooxygenase activity or platelet aggregation in human volunteers (Janssen et al., 1998)
Janssen et al., (1998) in ‘effects of flavonoid on hemostasis: Results from in vitro and a dietary supplement study wrote that only high concentrations of these flavonoids about 2500 µmol/L, which cannot be attained in-vivo by dietary consumption, inhibit collagen-induced aggregation in vitro. From their data, peak apigenin concentration in human plasma was < 1.1 µmol/L. The concentration we administered may have been enough to inhibit cyclooxygenase in relation to ovulation.
In conclusion,these studies suggest that administration of G. kola seed extract may partially block ovulation, alters oestrous cycle with a prolonged dioestrous, and may cause a dose dependent adverse effect on foetal development in S-D rats.









{kind=link}
{kind=link}
{kind=link}
I was diagnose with hpv virus for 5years i broke up with my boyfriend because i contracted the virus from him, luckily i found a herbalist name Dr. onokun everyone testify good of his natural treatment i picked up his address and contacted him. he truly treated me with natural herbs in a week and i got cured. i share this post because ever since i got this treatment I'm perfectly ok with my health. i contacted him through this email.. dronokunherbalcure@gmail.com
ReplyDelete